Poprzedni rozdział. | Wstęp do kognitywistyki - spis treści.
Informacja w mózgach: 14.1. Kora nowa | 14.2. Lokalizacja funkcji psychicznych | 14.3. Mapy topograficzne | 14.4. Kodowanie populacyjne.
Trzy podstawowe funkcje kory mózgu to:
- Analiza danych przychodzących ze zmysłów.
- Kontrola ruchów.
- Wyższe czynności psychiczne: planowanie, myślenie, analiza emocji.
Analiza sygnałów zmysłowych powinna z nich wydobyć wszelkie informacje przydatne do działania.
Dokonuje się to w kilku etapach.
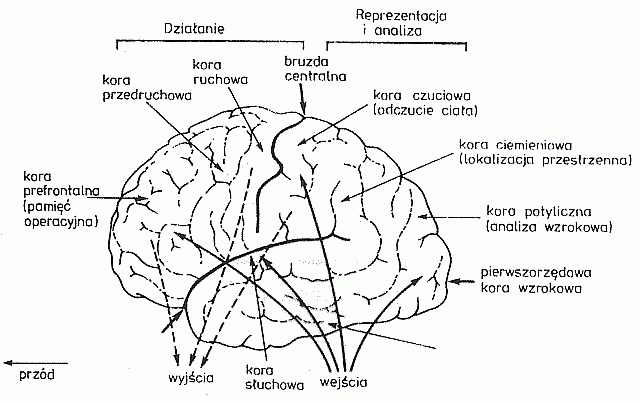
Pierwotne obszary kory otrzymują wstępnie przetworzoną informację od wzgórza i odpowiedzialne są za szczegółową analizę bodźców zmysłowych o określonej modalności. Skąd przychodzi, z jakiego miejsca ciała, z wnętrza czy skóry, oka czy ucha? Chociaż to seria impulsów to pobudzają się inne obszary kory, mają więc różną lokalizację w korze mózgu. Jakiego rodzaju receptory ją przysyłają? To pozwala rozróżnić, czy seria impulsów związana jest z temperaturą, ostrym bólem, czy głośnym dźwiękiem o określonej wysokości, barwie, konturze amplitudy lub kontraście widzianych krawędzi.
Wtórne obszary projekcyjno-kojarzeniowe przylegają do obszarów pierwszorzędowych i pozwalają na określenie znaczenia bodźców, interpretację sensu informacji. Mózg już wie, że dotarła do niego informacja z lewej stopy, czy to tylko ucisk czy ból bo coś ją przygniotło? Określenie kształtu czy kierunku ruchu wymaga połączenia informacji z wielu sąsiadujących receptorów.
Trzeciorzędowe obszary skojarzeniowe integrują informację z różnych zmysłów, pozwalając na rozpoznanie złożonych bodźców, są podstawą by utworzyć percepty, postrzeżenia lub wyobrażenia będące treścią naszego umysłu.
U makaka ponad 50% kory zajmuje się wzrokiem, 11% to kora somatosensoryczna, a tylko 3% kora słuchowa, chociaż dostarcza ona informacji do wielu obszarów mózgu. Kora wielu zwierząt zajmuje się przede wszystkim analizą danych zmysłowych i reakcjami ruchowymi na rozpoznane percepty.
W tej części popatrzymy na podstawowe mechanizmy, pozwalające na analizę informacji zmysłowych i kontrolę ruchu.
A14.1. Kora nowa
Receptory spełniły swoją funkcję i w mózgu pojawiają się różne impulsy. Zanim dotrą do świadomości przechodzą przez kilka obszarów mózgu, w których wydobywana jest z nich użyteczna informacja: w pierwotnej korze zmysłowej podstawowe cechy, takie jak z jakiej części ciała i jakiego rodzaju receptorów.
Obszary kory są w znacznym stopniu wyspecjalizowane; mózg nie jest uniwersalnym komputerem! Jednak kora potrafi się szybko dostosować do nowych bodźców, co widać w przypadku substytucji zmysłów lub uszkodzenia któregoś ze zmysłów lub części kory - inne obszary potrafią w pewnym stopniu się nauczyć nowych, przydatnych funkcji. Dlatego po udarze mózgu i zniszczeniu części kory możliwa jest rehabilitacja. Jest to jednak proces powolny i zwykle nie da się w pełni odzyskać pierwotnej sprawności. Kora różnych zwierząt przetwarza informacje z wielu typów receptorów, umożliwiając decyzję na najwyższym poziomie sterowania zachowaniem całego organizmu.
Chociaż zwykle usiłuje się przypisać funkcje wyróżnionym obszarom kory na podstawie reakcji na określone bodźce lub zadania, lepszym podejściem jest analiza rodzaju przetwarzania informacji, transformacji sygnałów dokonywanej przez dany obszar. Można to wywnioskować na podstawie analizy aktywności danego obszaru mózgu w różnych zadaniach.
Kora nowa (łać. cortex = kora,
neocortex) jest odpowiedzialna za złożone czynności poznawcze.
Tylne i boczne części kory mózgu specjalizują się w analizie danych zmysłowych. Część najbardziej wysunięta do przodu, czyli kora przedczołowa, zajmuje się planowaniem działań, przechowuje informacje w pamięci roboczej i wydaje polecenia korze czołowej, która wspólnie z dużymi jądrami podstawy mózgu
(ciałem prążkowanym)
przygotowuje sygnały przesyłane do móżdżku, rdzenia kręgowego i na końcu do mięśni.
Kora nowa człowieka zawiera około 14-18 mld neuronów i około 60 mld komórek glejowych. Chociaż to zaledwie 1/5 wszystkich neuronów w mózgu to są to duże neurony.
Jeśli uwzględnić połączenia, białą materię biegnącą przez głębsze obszary, to kora zajmuje około 80% objętości mózgu, a jej masa to również około 80% masy całego mózgu. Widać więc, jak ważne są połączenia, czyli konektom.
Kora ma grubość od 1.5 mm (kora wzrokowa) do 4.5 mm (kora ruchowa), średnio ok 3 mm, w pionowym przekroju w jednej linii znajdziemy ok. 50-100 neuronów. Widoczna jest tylko 1/3 na powierzchni bocznej, pozostała część kory jest na powierzchni przyśrodkowej i podstawnej.
Jej powierzchnia jest trudna do zmierzenia ze względu na silne pofałdowanie; wymiar fraktalny kory to ok. 2.8, więc bardzo wysoki, jest obiektem prawie wypełniającym przestrzeń w trzech wymiarach, a nie płaską, dwuwymiarową warstwą. Nowsze źródła
(Human Connectome Project) podają prawie 1000 cm2 w każdej półkuli, czyli wielkość średniej pizzy. Średnia wartość jest oceniana na około 1 840 ± 200 cm2
(Van Essen, Donahue & Glasser, 2018).
|
Są 3 typy kory:
- Izokora, filogenetycznie młoda, ma 6 warstw (większość kory).
- Allokora - paleokora (np. kora węchowa) i archeokora (np. kora hipokampa), stara, 3 warstwy.
- Mezokora - kora pośrednia (np. w zakręcie obręczy, okolicach hipokampa), warstwy 2,3,4 zlewają się w jedną warstwę.
Neurony są zróżnicowane: dwie ważne kategorie to neurony wstawkowe (interneurony, o krótkich aksonach, ok. 20%) i neurony projekcyjne, które mają długie aksony (jest ich ok 80%), takie jak
neurony piramidalne.
Kora ma budowę modularną
(dokładniejsze informacje), można w niej wyróżnić zarówno warstwy jak i kolumny korowe.
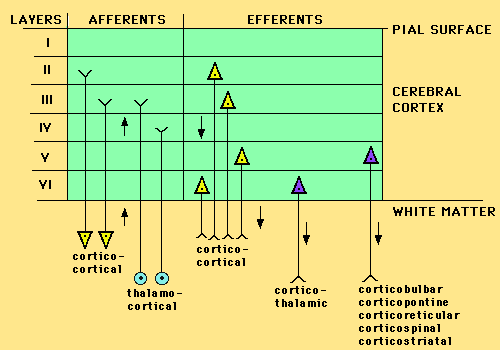
Budowa laminarna kory to 3 (stara allokora) lub 6 warstw (izokora) o specyficznej organizacji. Warstwy 2-4 otrzymują pobudzenia z innych części kory i wzgórze (aferentne, czyli wchodzące), warstwy 2, 3, 5, 6, wysyłają swoje aksony do kory, wzgórza i pnia mózgu (to połączenia projekcyjne, lub eferentne). Warstwa 1 nazywana "molekularną" zawiera niewiele ciał neuronów.
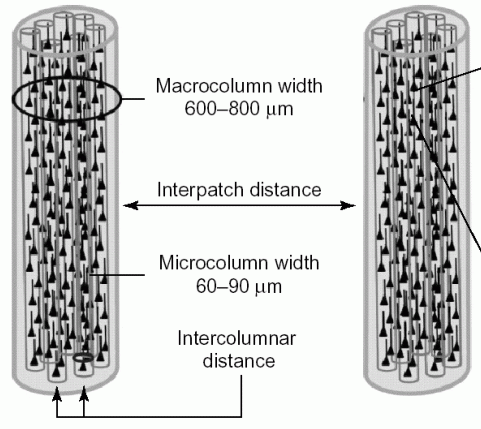
Gęste połączenia neuronów w pionie a rzadsze w poziomie pozwalają wyróżnić
minikolumny, które mają zwykle 80-120 neuronów, dwa razy tyle w korze wzrokowej.
Kolumny korowe, lub makrokolumny są znacznie większe, mają 10-100 tysięcy neuronów, czyli 70-1000 minikolumn. Łatwiej jest je odróżnić i czasami przypisać konkretną funkcję, np. każdy wibrys (włos wąsa szczura) jest połączony z jedną kolumną analizującą dochodzące z niego wibracje.
|
M. A. Hofman
uważa, że w
minikolumnie korowej
jest 108 neuronów a makrokolumna, którą można uznać za w miarę odrębny moduł kory, ma tyle minikolumn ile jest neuronów projekcyjnych z niej wychodzących; u człowieka jest ok. 70 minikolumn, czyli ok. 7600 neuronów w module, każdy połączony z ok. 1000 innych, w sumie około 3 mln takich modułów. Nie są to jednak dokładne oceny, wielkość kolumn i grubość kory jest zróżnicowana.
Chociaż liczba takich kolumn jest znacznie mniejsza niż liczba pojedynczych neuronów, to liczba możliwych stanów wewnątrz nich (czyli oscylacji lokalnych pętli znajdujących się w kolumnach mikroobwodów neuronów) jest praktycznie nieskończona.
Teoria takich modułów i próba ustalenia jakie dokładnie pełnią funkcje jest opracowywana w ramach kognitywnych neuronauk obliczeniowych (np. Rinkus, 2010).
Biała materia to długie aksony neuronów łączących odległe obszary mózgu: jest ich około 3 mld, długość sumaryczna 150.000 km, ok. 200 mln takich aksonów tworzy spoidło wielkie. Biały kolor ma
otoczka mielinowa aksonu, substancja chroniąca i izolująca elektrycznie włókno nerwowe, dzięki czemu impulsy elektryczne nie tracą swojej energii. Z gęstością większą niż 1 mm w tej osłonce są niewielkie
przewężenia Ranviera (rzędu 1/1000 mm), dzięki którym sygnał jest dodatkowo wzmacniany.
Szybkość rozchodzenia się sygnałów przez długie aksony to ok. 10 m/sek, czyli w 20 ms informacja może się rozejść po całym mózgu.
Szybkość przesyłania impulsów
różni się w zależności od typu neuronów, najszybciej działają
motoneurony alfa (do 120 m/s), wrażenia dotyku i ostrego bólu ponad 70 m/s, najwolniej reagują niektóre neurony bólowe (0.5 m/s).
Najwięcej białej materii i najszybsze czasy reakcji mamy około 40 roku życia.
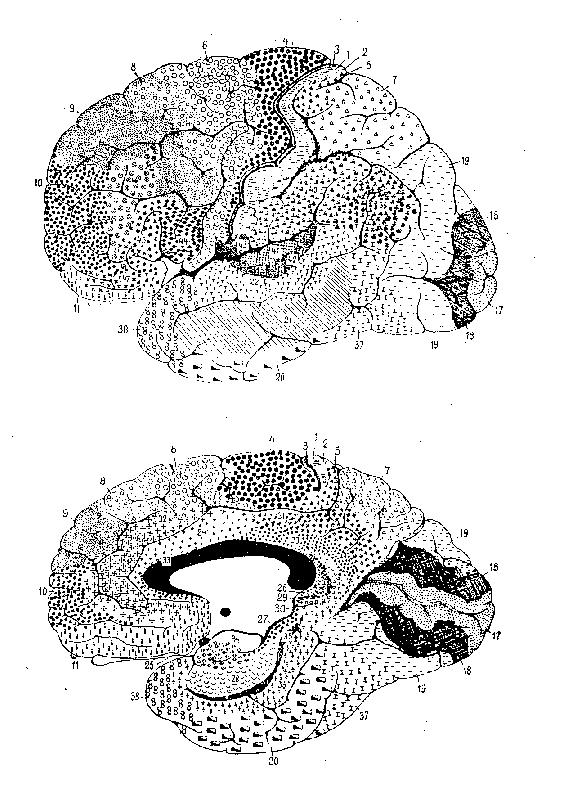
52 cytoarchitektonicznie jednorodne obszary kory nazywają się
polami Brodmanna (1909 rok). Taka parcelacja kory mózgu jest nadal powszechnie używana.
Interaktywny atlas
podsumowujący różnorodne funkcje związane z aktywnością wybranego pola został zrobiony na podstawie badan fMRI,
oraz materiały do nauki
neuroanatomii KenHub .
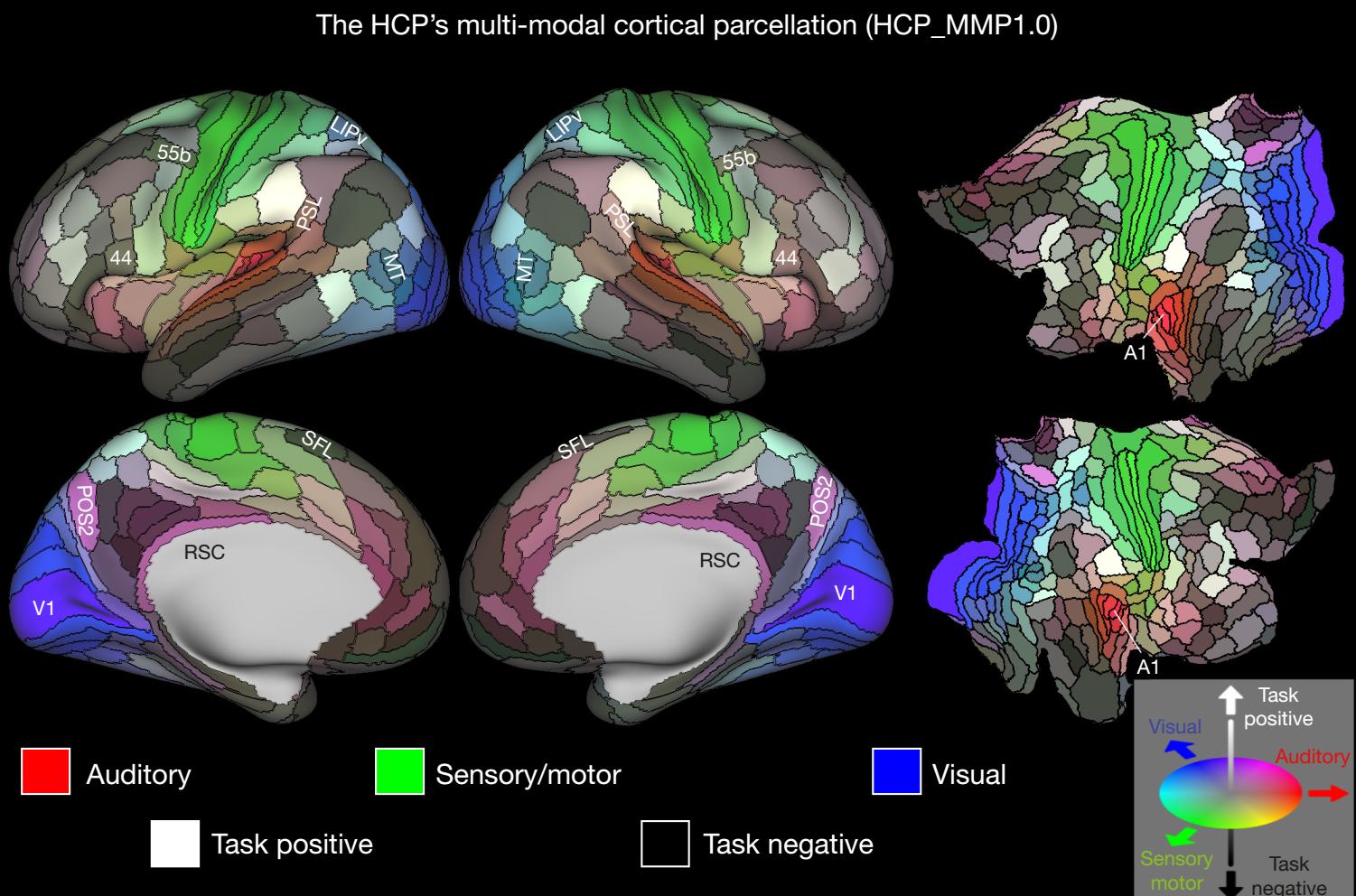
Nowsze podziały były mniej popularne, ale w 2016 roku pojawiła się znacznie lepsza parcelacja kory na 180 obszarów w każdej z półkul, która powinna stać się standardem. Znana jako HCP’s multi-modal parcellation, v1.0 (Glasser i inn, 2016), opracowana została wykorzystując metody konektomiki opracowane w ramach
Human Connectome Project.
Jest to uśredniony model kory oparty na danych ponad 200 zdrowych osób. Każdy wyróżniony region może się znacznie różnić u indywidualnych osób, grubość kory różni się nawet dwukrotnie, a powierzchnia regionów to od 0.2-20 cm2.
{kind=link}
wzrokiem i podejmowaniem działań. |
Móżdżek zawiera 80% wszystkich neuronów, ale zajmuje tylko 10% objętości i masy mózgowia, a jego kora ma powierzchnię równą połowie kory nowej i grubość ok. 1 mm, a więc neurony są w nim mniejsze i znacznie gęściej upakowane.
Ośrodki podkorowe zajmują poniżej 10% objętości mózgowia i zawierają niecały 1% wszystkich neuronów. Zgrupowane są w setkach małych jąder i pełnią bardzo ważną rolę.
Są to duże neurony mające rozległe połączenia (biała materia), zanurzone w gęstej materii dendrytów, aksonów, komórek gleju i naczyń krwionośnych, nazywanej
neuropilem.
Organizacja strukturalnych sieci jest widoczna na obrazach tworzonych techniką DTI, czyli diffusion tensor imaging fMRI. Takie obrazy pokazują przebieg długi wiązek aksonów neuronów, łączących odległe obszary mózgu. Kolory są sztucznie nadawane, zależnie od kierunku połączeń, zwykle lewa-prawa, przód-tył, góra-dół. Czasami (jak na rysunku poniżej) kolor oznacza połączenia pomiędzy podobnymi obszarami kory i podkorowymi strukturami.
Badając najbardziej gęste wiązki połączeń można zidentyfikować główne węzły połączeń (huby) i przedstawić uproszczoną strukturę sieci w postaci grafu. Wielkość węzłów na grafie zależy od liczby połączeń danego obszaru. Na rysunku widać największe węzły w obszarach kory czołowej, ciemieniowej, okolicach hipokampa, wzgórza i skorupy (jądra podstawy).
Badając zmianę sygnału w czasie można zauważyć obszary mózgu zachowujące się w podobny sposób, zmieniające swoją aktywność w synchroniczny sposób, a więc powiązane funkcjonalnie. Na kolejnych rysunkach obszary analizowane są z coraz większą dokładnością, ukazując grupy neuronów wewnątrz modułów, realizujących określone funkcje.
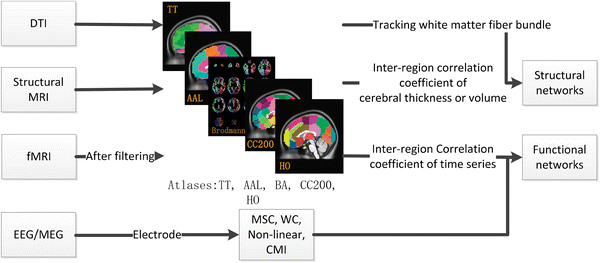
funkcjonalne fMRI dla aktywacji, EEG/MEG dla analizy szybkich zmian. Rys. z Structural and Functional Brain Networks, 2013 |
Główne typy neuronów i ich połączenia są z grubsza znane, ale jest wiele rzadkich typów neuronów i nie wszystkie zostały już opisane. Połączenia pomiędzy neuronami w mózgu dzieli się na kilka sposobów:
- aferente lub eferentne;
- projekcje, pęczki, spoidła;
- ruchowe, czuciowe lub mieszane;
- czaszkowe lub rdzeniowe;
- korowo-korowe, korowo-wzgórzowe lub korowo-limbiczne;
- specyficzne (niosące określone informacje do określonych obszarów kory, np. z receptorów zmysłowych)
- niespecyficzne (zwykle rozproszone, wywołujące ogólne pobudzenie, lub niosące informację o różnej modalności).
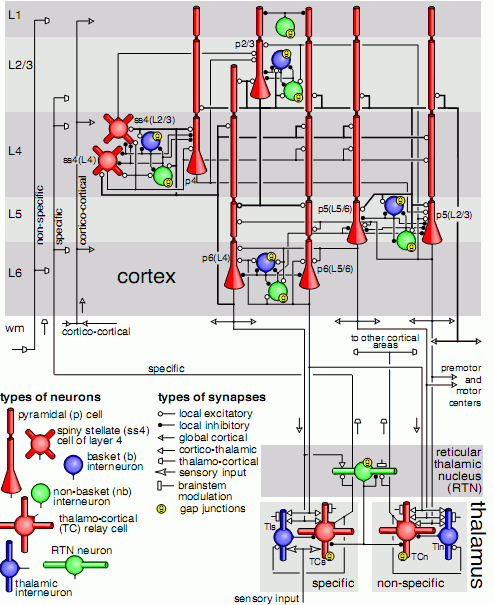
Large-scale model of mammalian thalamocortical systems. |
Szczegółowy obraz kory na poziomie pojedynczych neuronów jest bardzo skomplikowany, struktura połączeń jest nieregularna. Jest wiele typów neuronów, synaps, kanałów jonowych reagujących w różny sposób na neurotransmitery, ale ogólny schemat połączeń jest podobny w różnych częściach kory. W tym szaleństwie jest jednak metoda: neurony połączone w mikroobwody reagują oscylacjami, które mają różny okres trwania i różne częstotliwości. Pozwala to zamienić serię impulsów dobiegających z różnych źródeł na sygnał łatwiejszy do odczytania i trwający nieco dłużej niż samo pobudzenie zmysłów. Trzask gałęzi może być bardzo krótki, ale w mózgu wywoła wibracje mikroobwodów, które zostaną przeanalizowane, sieć uwagowa zorientuje nas w stronę hałasu i przygotuje do szybkiego działania. Może to skradający się drapieżnik ...
Na rysunku kolorem czerwony zaznaczone są neurony pobudzające, a niebieskim interneurony hamujące o krótkich aksonach. Schemat połączeń jest na tyle dobrze poznany, że możliwa jest symulacja komputerowa fragmentów kory.
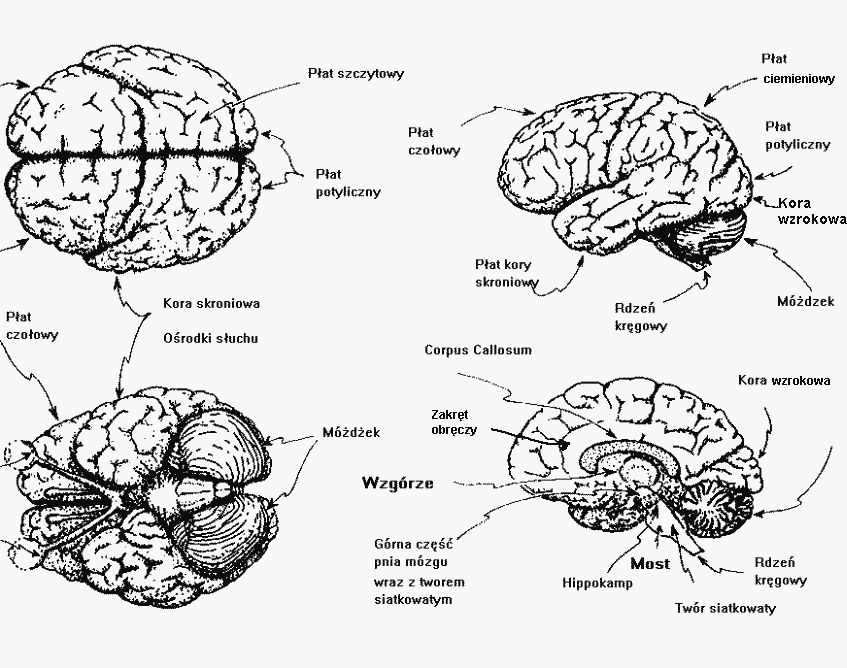
Kora podzielona jest na dwie półkule, 5 płatów, liczne zakręty i bruzdy.
Szkice na filmach ilustrujące wygląd kory (animacje z
Digital Anatomist Project):
2-minute neuroscience videos, cerebral cortex, prefrontal cortex, limbic system.
Large-scale model of mammalian thalamocortical systems. |
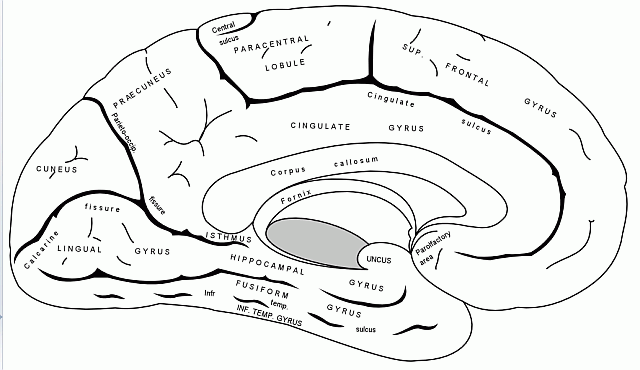
Płat limbiczny jest ukryty pomiędzy półkulami i można go podzielić na wiele obszarów o różnych funkcjach.
|
Uszkodzenia rozwoju kory prowadzą do ciężkich upośledzeń.
Mity miejskie: pewien szkocki matematyk o wysokim IQ prawie nie miał kory mózgu. Okazało się to pomyłką lekarza, który źle opisał rentgenograficzne zdjęcia.
Drugim źródłem jest doniesienie o obserwacjach J. Lorbera w Science 210 (1980), ale również
nie potwierdzone, a zapowiadana praca naukowa nie została nigdy opublikowana. Jest też kilka innych doniesień o poważnym wodogłowiu, ale brakuje badań pokazujących ile i w jakim stanie zostało kory mózgu i jak to się odbiło na działaniu mózgu. Przypadek chłopca Noah Wall
opisany został w 2018 roku w czasopismie
NeuroQuantology, nie uznawanym za naukowe
(niestety przedrukowali jeden z moich artykułów z 2002 roku).
Dwóch autorów, z których żadne nie pracuje w naukowej instytucji, próbowało dokonać neuro-kwantowej interpretacji medycznego cudu. Podchwyciła to po latach popularna prasa donosząc, że chłopiec urodził się mając 2% mózgu (nie wiadomo jednak, co to ma znaczyć). Ich rewelacje pochodzą nie od lekarzy, tylko od rodziców chłopca, który urodził się z wodogłowiem. Autorzy twierdzą, że to wola by przeżyć spowodowała, że chłopcu przybyło 80% normalnego mózgu. Żaden lekarz czy neuronaukowiec nie zainteresował się takim cudem. Za to rodzice zarabiają na popularności chłopaka.
Szybkie wnioski: bez kory umysł też istnieje! Myśli dusza!
Prawda (smutna?): jest wyraźna
korelacja stopnia uszkodzenia kory i upośledzeń poznawczych i ruchowych.
Duża plastyczność kory pozwala przejąć jej pewne funkcje (zwykle po długotrwałym treningu) jeśli uszkodzenia nastąpiły we wczesnym okresie rozwoju, w późniejszym okresie możliwości kompensacji są znacznie mniejsze.
Czy kora jest konieczna do normalnego działania organizmu?
Całkowite bezmózgowie kończy się śmiercią, ale mając bardzo rzadką wrodzoną chorobę, jaką jest
hydranencefalia,
czyli brak większości normalnej kory, można przy odpowiedniej opiece przeżyć (najstarsza osoba zmarła w wieku 32 lat), chociaż większość umiera w pierwszych latach życia.
Co ciekawe, pomimo wielkich problemów rozwojowych takie osoby zachowują się w sposób "jakby świadomy", a przynajmniej przytomny, orientują się w środowisku (głównie za pomocą słuchu), okazują emocje, ich zachowanie jest typowe dla ludzi.
Takie obserwacje są zgodne z przekonaniem, że większość odruchów typowych dla danego gatunku jest rezultatem działania ośrodków podkorowych (limbicznych i pnia mózgu). Jednakże nie zawsze są to osoby dokładnie zdiagnozowane, część może mieć szczątkową korę. Bez niej nie ma możliwości autorefleksji, a więc i samoświadomości, ale też bardziej precyzyjnego działania, percepcji, czy planowania.
Gryzonie i koty pozbawione kory zachowują się prawie normalnie, wykazując tylko niewielkie zmiany zachowania
(Merker 2006).
A14.2. Lokalizacja niektórych czynności psychicznych
Początkowo uważano, że w mózgu nie ma wyróżnionych obszarów realizujących wyspecjalizowane funkcje.
|
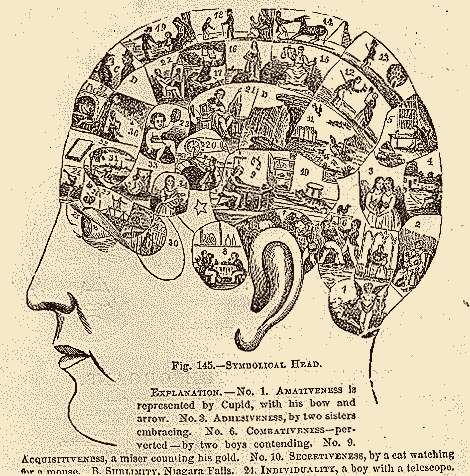
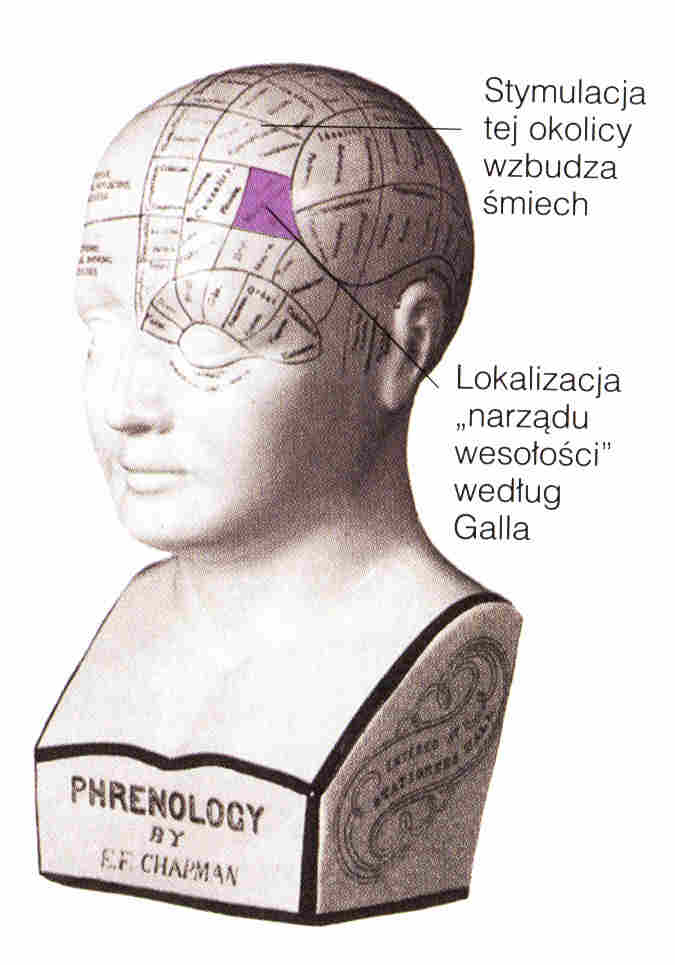
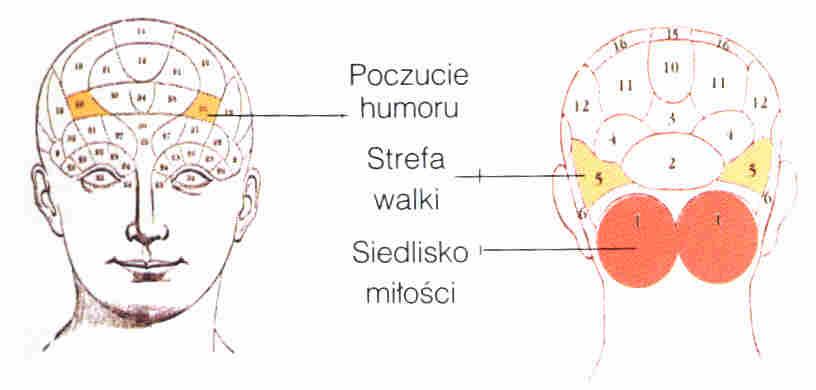
Lokalizacja funkcji: frenologia (kranioskopia), w XIX wieku niezwykle popularna pseudonauka.
"Odkryto" 37 obszarów: skłonności do stałości, ostrożności, duchowości, kochliwości, opiekuńczości, zdolności językowych... Franz Joseph Gall i Johann Spurzheim zebrali tysiące obserwacji potwierdzających ich system! Był to pierwszy krok w stronę lokalizacji funkcji i okazuje się, że drażnienie kory niedaleko od podanego przez nich obszaru humoru wywołuje śmiech. Oczywiście nie jest to "region śmiechu", ale kluczowy obszar pobudzający odpowiednie sieci. Początkowo odrzucano "lokalizację ducha", ale frenologia stała się niezwykle popularna i w postaci psychognomiki Bouta spotyka się ją do dziś! Z Chin i Bliskiego Wschodu wywodzi się pokrewna pseudonauka,
fizjonomika, odczytująca charakter z kształtu twarzy.
|
{kind=link}
|
|
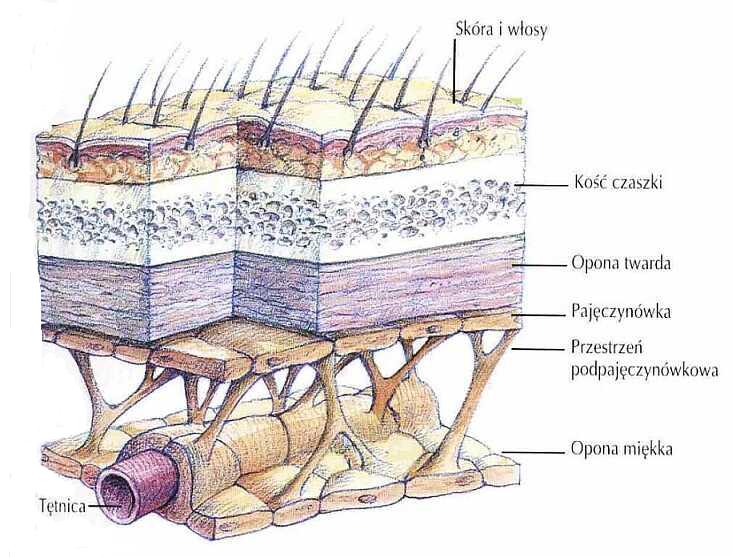
Frenologia nie ma szans by coś powiedzieć o mózgu, bo jest on dobrze chroniony przez kilka warstw pomiędzy kośćmi czaszki i korą. Opony mózgu i pajęczynówka to swojego rodzaju amortyzacja.
Płyn mózgowo-rdzeniowy mieści się w przestrzeni między oponami i w komorach mózgu.
Kształt mózgu nie uwidacznia się na powierzchni czaszki. Pewne znaczenie ma grubość kory, ale nie zostawia ona śladów na powierzchni czaszki.
Odległe pobudzenia dochodzące do kory (aferentne):
|
- Włókna kojarzeniowe z tej samej półkuli (pęczki, obręcze).
- Włókna kojarzeniowe przeciwległej półkuli (spoidła).
- Projekcje z wzgórza.
- Projekcje z pnia mózgu.
Odległe pobudzenia wychodzące z kory (eferentne):
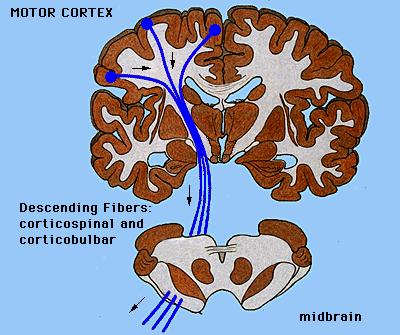
|
- Włókna kojarzeniowe z tej samej półkuli (pęczki, obręcze).
- Włókna kojarzeniowe przeciwległej półkuli (spoidła).
- Projekcje sensomotoryczne, skojarzeniowe i limbiczne do prążkowia.
- Projekcje ruchowe do nerwów czaszkowych, pnia mózgu
(droga korowo-mostowa, korowo-siatkowa) i rdzenia.
Przekrój pokazujący spoidło łączące obie półkule mózgu.
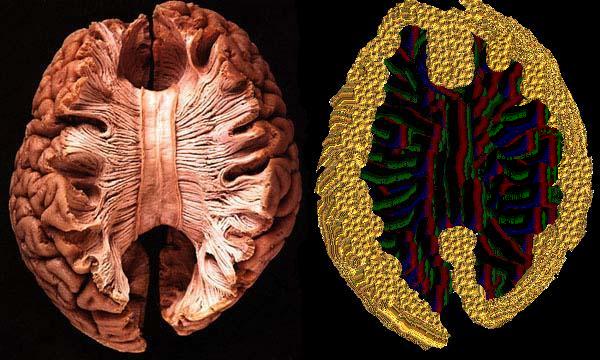{kind=link}
Postrzeganie wyrazu twarzy jest dla ludzi i małp naczelnych bardzo ważne. U człowieka powstały dwa odrębne mechanizmy sterowania mięśniami twarzy (Gazzaniga i Smylie, 1999, skąd pochodzą rysunki poniżej).
Mięśnie górnej i dolnej części twarzy kontrolowane są w różny sposób.
Górna i dolna część twarzy sterowana jest przez prążkowie, czyli podkorowe jądra zaangażowane w ruchy automatyczne
(to układ pozapiramidowy).
Mięśnie wokół ust i poniżej sterowane są dodatkowo przez pierwszorzędową korę ruchową (to
układ nazywany piramidowym). Dzięki temu człowiek zdolny jest do ukrywania uczuć.
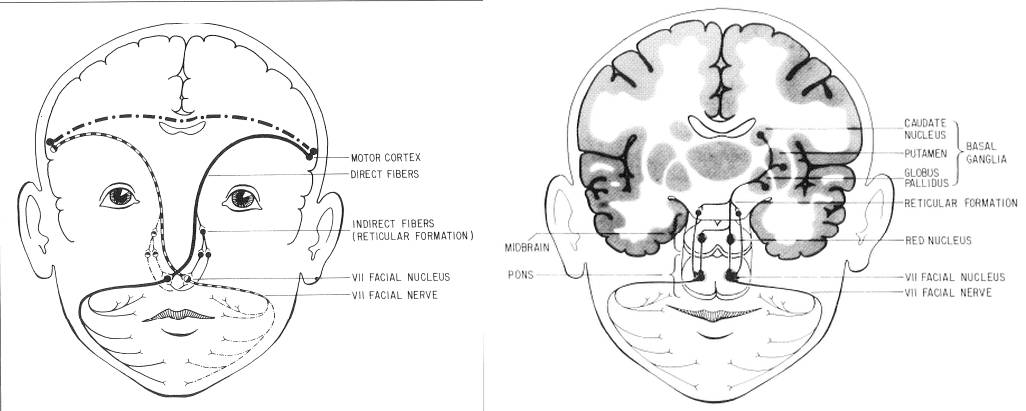
|
A14.3. Mapy topograficzne
Neurofizjolodzy podzielili korę na 3 rodzaje: związaną ze zmysłami, ruchem i skojarzeniami. Każdy z tych obszarów wykorzystuje odmienne mechanizmy przetwarzania informacji. Jest to:
- mapowanie topograficzne;
- kodowanie populacyjne;
- mechanizmy skojarzeniowe.
Analiza sygnałów zmysłów: najpierw odwzorowanie na powierzchni kory przy zachowaniu relacji topograficznych i redukcji wymiarowości.
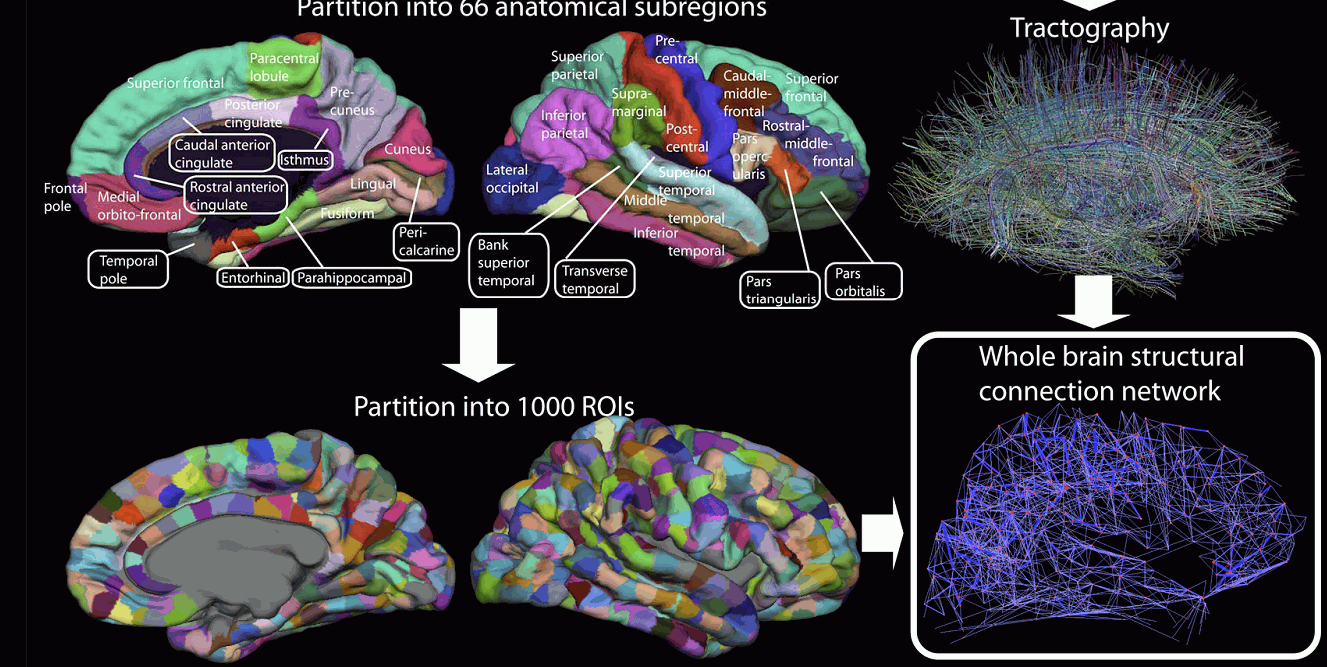
Bardziej szczegółowa analiza kory (w pracy Human Connectome Project multi-modal parcellation, Glasser i inn, 2016), opracowana została wykorzystując metody konektomiki w ramach
Human Connectome Project, po uśrednieniu wyników dla 210 zdrowych osób.
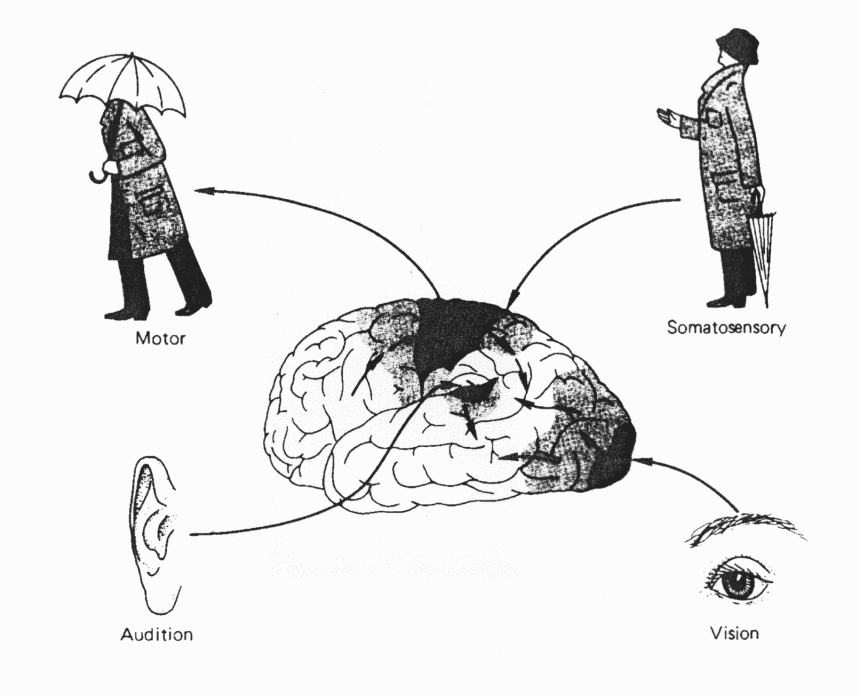
Przydałby się dobry schemat mózgu na rożnych poziomach szczegółowości.
Poniżej jest tylko stary szkic, z zaznaczeniem głównych struktur i szlaków je łączących przy wykorzystaniu różnych neurotransmiterów:
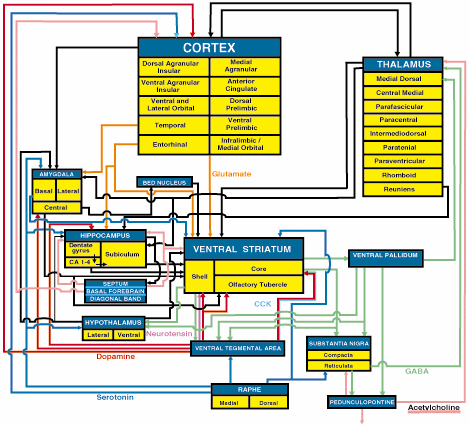
Dwie główne struktury to kora nowa (wyróżniono korę wyspy) jak i wzgórze. Widzimy też ważne jądra podkorowe: prążkowie brzuszne (Ventral striatum), hipokamp, ciało migdałowate, oraz podwzgórze, oraz kilka jąder śródmózgowia (substancja czarna) i pnia mózgu (jądra szwu). Schemat ten jest dalece niekompletny.
"Mapa umysłu" podsumowująca
funkcje płatów kory.
Podział kory na obszary o podobnej budowie komórkowej nie wystarcza do zrozumienia jej funkcji. Najważniejsze są podsieci łączące ze sobą różne obszary.
Dzięki traktografii można stworzyć mapę połączeń pomiędzy wszystkimi regionami mózgu, czyli opisać konektom strukturalny.
Uczenie się jest związane ze zmianą struktury połączeń, ale ta sama struktura fizyczna może realizować różne procesy, podobnie jak się to dzieje w komputerze, który nie zmieniając swojej struktury realizuje całkiem inne programy. Dlatego istotne jest rozróżnienie połączeń strukturalnych, które tworzą konektom, od połączeń funkcjonalnych, które stanowią podzbiór połączeń konektomu przez który przepływa informacja przy wykonywaniu konkretnego zadania.
W ogromnej sieci mózgu rozchodzenie się aktywacji musi zachodzić w precyzyjnie określonych podsieciach, w przeciwnym przypadku energia rozpłynie się wszystkimi ścieżkami i nie będzie można rozpoznać, w jakim stanie znajduje się mózg, jaka myśl lub jaki percept pojawia się w danym momencie. Można taką sytuację przyrównać do zbyt słabego kontrastu obrazu, na którym trudno wyróżnić jakieś kształty i rozpoznać obiekty.
Im bardziej szczegółowa jest parcelacja obszarów mózgu i dokładniejszy konektom tym więcej można zebrać informacji o specyficznych podsieciach zaangażowanych w realizację określonego zadania.
Struktura konektomu ustala się dopiero w wieku dojrzałym.
Połączenia strukturalne zmieniają się w miarę uczenia się powoli dzięki neuroplastyczności, zmianom synaptycznym.
Połączenia funkcjonalne to podstawa do realizacji procesów dynamicznych, które możemy powiązać z percepcją i procesami mentalnymi. Konfiguracje sieci funkcjonalnych zmieniają się bardzo szybko gdy przerzucamy się pomiędzy wykonywanymi zadaniami. Zmieniają się również na skutek ogólnego pobudzenia mózgu, np. silnych emocji, kofeiny, pragnienia czy głodu.
Torowanie (prymowanie) w wyniku podawania jakiejś wskazówki czy bodźca ułatwia pobudzenie specyficznych sieci funkcjonalnych, a więc przypomnienie sobie lub szybsze skojarzenie w późniejszym okresie. Na poziomie neuronalnym wiąże się to prawdopodobnie z obniżeniem progu pobudzenia neuronów należących do wybranej podsieci.
Podział na choroby psychiczne, zaburzenia psychiatryczne i neurologiczne nie jest całkiem jednoznaczny. Z grubsza można uznać następujące rozróżnienie.
- Zaburzenia psychiczne (np. nerwice) dotyczą procesów mentalnych, funkcjonowania mózgu na poziomie dynamiki sieci funkcjonalnych. Psychoterapia może doprowadzić poprzez pobudzenia emocji lub nawet rozmowę do zmiany zachowania aktywizując inne podsieci.
- W przypadku zaburzeń psychiatrycznych zmiany są zwykle głębsze i dotyczą braku biochemicznej równowagi lub niewłaściwego funkcjonowania neuronów (np. depresja, choroba Alzheimera), można je więc do pewnego stopnia leczyć farmakologicznie.
- Zaburzenia neurologiczne są wynikiem uszkodzeń układu nerwowego, np. uszkodzeń naczyń krwionośnych, udaru lub uszkodzeń nerwów obwodowych.
Czucie powierzchniowe i kontrola ruchów oparte są o podobne mechanizmy, to: mapy somatosensoryczne i mapy ruchowe.
Mapy somatosensoryczne opisał w 1951 roku neurochirurg
Wilder Penfield, który stymulował prądem różne obszary kory osób cierpiących na padaczkę przez operacją (to była standardowa procedura by określić, czy nie uszkodzi się istotnych funkcji), notując wrażenia, jakie taka stymulacja powodowała.
Wielkość obszaru analizującego dane zmysłowe zależy od ich przydatności dla danego zwierzęcia, stąd duże różnice w powierzchni kory przeznaczonej do analizy różnych sygnałów.
Gryzonie mają dużą powierzchnię poświęconą wibrysom. U człowieka dominuje reprezentacja rąk i języka.
Informacja somatosensoryczna (czucie ciała) odbierana jest przez obszar SI kory, znajdujący się z tyłu bruzdy centralnej.
Informacja przekazywana jest od receptorów: dotyku, bólu, temperatury, wibracji, położenia kończyn, przez nerwy czuciowe do wzgórza i kory SI, w której różne grupy neuronów specjalizują się w analizie różnego typu informacji.
Szlaki czuciowe (film tylko lokalnie)
(animacja z Digital Anatomist Project).
Różne pobudzenia docierają różnymi nerwami: nerwy przekazujące sygnały szybko sygnalizują ból ostry, a wolno ból piekący. Blokowanie przepływu różnych sygnałów może nastąpić już w rdzeniu kręgowym, dlatego ucisk pomaga zmniejszyć wrażenie bólu.
Korelacje ruchu i dotyku: pobudzenia z przeciwległych obszarów kory SI dochodzą do pobliskich obszarów MI (za bruzdą centralną). Dzięki temu możliwe są szybkie reakcje na bodźce dotykowe (działania sensomotoryczne), np. odsunięcie ręki przy ukłuciu czy oparzeniu.
Twarz i język reprezentowane są częściowo po tych samych stronach, a pozostałe części ciała przeciwlegle.
Zniszczenie kory SI powoduje zanik wrażeń czuciowych, jednak ból i temperatura po pewnym czasie pojawiają się; zgrubne rozróżnianie tych wrażeń możliwe jest już na poziomie wzgórza.
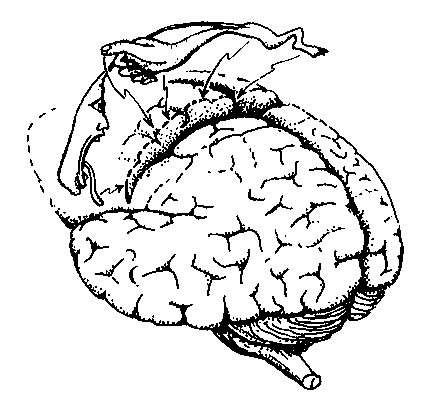
Różne części wzgórza zaangażowane są w projekcje somatosensoryczne:
VPL=Ventral Posterior Lateral Nucleus = jądro brzuszno-tylne boczne.
VPM = Ventral Posterior Medial Nucleus = jądro brzuszno-tylne przyśrodkowe.
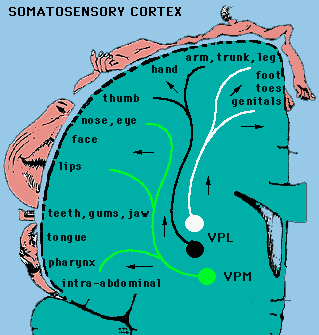
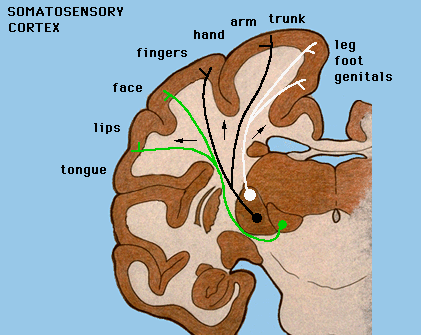
Mapy czuciowe: pobudzanie kory czuciowej SI słabym prądem wywołuje wrażenia dotyku, łaskotania, swędzenia, umiejscowiane w określonym obszarze ciała.
Pobudzanie kory MI z przodu bruzdy centralnej wywołuje zachowania ruchowe (całe wyuczone ruchy).
Są drobne różnice pomiędzy mapą ruchową i czuciową.
Obszary ciała mają podobne odwzorowanie w zakrętach kory, ale poza tym mamy duże indywidualne różnice i plastyczność (zmienność w czasie).
Kora SI ma budowę kolumnową (baryłkową) o średnicy rzędu ułamka milimetra: na pobudzenia reaguje cała kolumna.
Kora SI ma 4 podobszary Brodmanna:
- obszar 2 i 3a analizuje sygnały proprioceptywne, reakcje na ruch w określonym kierunku;
- obszar 3b reaguje na bodźce dotykowe
- obszar 1 reaguje na sygnały mieszane
Obszar kory poświęcony analizie jest proporcjonalny do wagi bodźców. Celem analizy jest precyzyjna dyskryminacja.
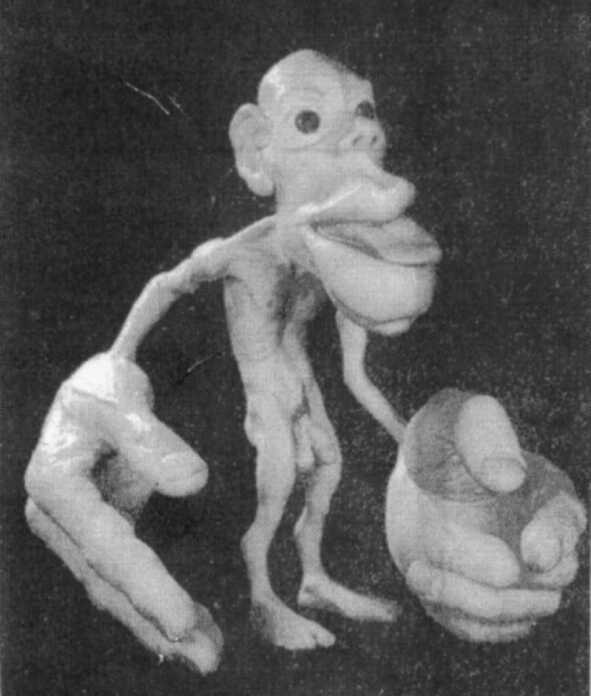

Homunculus Mapper, eksperyment zbierający dane na temat gęstości receptorów by zamienić je na figurkę homunkulusa. Podobny eksperyment przedstawiłem tutaj.
Zrób sobie mapę kory motorycznej
Mapa własnej skóry: trzeba sprawdzić czy czujemy różnicę przy dotykaniu jednego lub dwóch ostrych czubków (np. gwoździ czy ołówków), których wzajemną odległość możemy regulować. Jeśli ktoś będzie nas dotykał na zmianę, raz jednym raz dwoma ostrzami, zwiększając między nimi odległość, zaczniemy odczuwać różnicę; minimalna odległość na dłoni to milimetr, ale na plecach to kilka centymetrów.
Dotykając twarzy lokalizujemy odczucie dotyku na twarzy, a pleców bardziej ręki. Zależy to nie tylko od gęstości receptorów, ale od tego na czym skupiamy uwagę.
Plastyczność map czuciowych: do lat 1970 sądzono, że mózg nie zmienia się po osiągnięciu dojrzałości.
Paul Bach-y-Rita
prowadził pionierskie prace już w latach 1960, pokazując jak można doprowadzić do rehabilitacji mózgu po udarze.
Doświadczenia z małpami prowadzone były przez
Michaela Merzenicha oraz
Edwarda Tauba.
Metoda
wymuszania ruchu przy ograniczaniu (Constraint-induced movement therapy) opracowana przez Tauba przynosi bardzo dobre rezultaty.
Kryminalna historia Silver Spring Monkeys warta jest zastanowienia: miłośnicy zwierząt w dobrej wierze doprowadzili do zniszczenia dobrego laboratorium, opóźniając prace nad bardzo ważną metodą terapii, obecnie powszechnie stosowaną.
Historia rozwoju idei neuroplastyczności opisana jest w znakomitej książce N. Doidge: The brain that changes itself. Tu jest krótki film ilustrujący neuroplastyczność.
Zmiana wielkości obszaru kory reagującej na dane bodźce następuje na skutek stymulacji lub braku bodźców (po uszkodzeniu nerwu lub amputacji palca). Związanie kilku palców tak, że mogą robić ruchy tylko jednocześnie, powoduje powstanie jednej rozległej mapy.
Reorganizacja map czuciowych i ruchowych po uszkodzeniu i zrośnięciu nerwów przebiega w podobny sposób (rys. poniżej). Symulacje rozwoju map topograficznych po urazach mogą być przydatne w rehabilitacji.
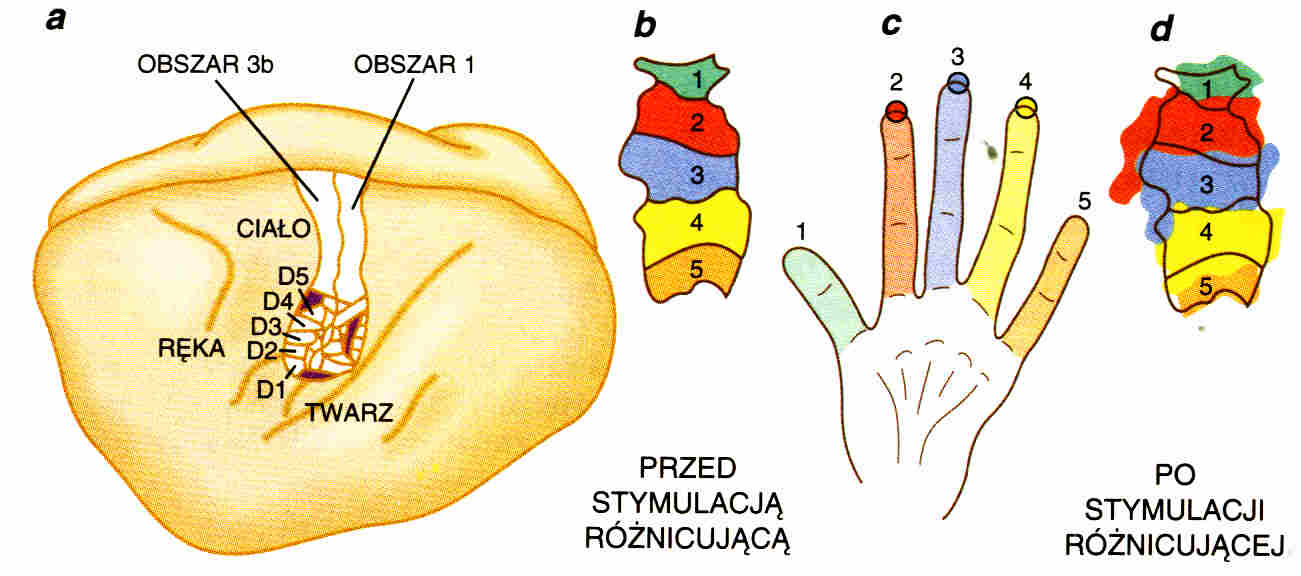
|
Przykład plastyczności: uczenie się rozpoznawania palców u nóg.
Czy naprawdę wiemy, co czujemy? Działanie kory czuciowej nie zawsze jest precyzyjne: który palec u nogi został dotknięty? Jeśli go nie widzimy dokładność odpowiedzi osiąga 80-90%. Korelacja wzrok-dotyk szybko podnosi dokładność do 98%. Chodzenie w butach po paru dniach obniży dokładność do początkowej. Bez dyskryminacji nie potrafimy określić swoich wrażeń - jest to podstawa świadomej percepcji. |
 |
Dylemat plastyczności-stabilności pojawia się tu kolejny raz.
Za duża stabilność reakcji kory => brak adaptacji, mózg się nie uczy.
Za duża plastyczność kory => możliwe jest katastroficzne zapominanie ważnych reakcji na istotne bodźce.
Konieczny jest kompromis na każdym poziomie!
- Neurony: kompromisu wymaga stabilność synaps i zmiany umożliwiające uczenie się reakcji neuronów na nowe bodźce.
- Mózg: kompromisu wymaga uczenie się nowych faktów i jednoczesne zachowanie stabilnego, realistycznego obrazu świata.
- Mózg się częściowo sam naprawia dzięki neuroplastyczności, ale liczne uszkodzenia prowadzą do szybkiej śmierci (np. w obrębie pnia mózgu, regulującego oddychanie i inne funkcje homeostatyczne).
- Zmiany genetyczne mogą uszkodzić też komórki macierzyste i tworzyć wadliwe komórki.
- Społeczeństwa: kompromisu wymaga stabilność (tradycja, partie konserwatywne, jeśli coś działa lepiej nie zmieniać) i konieczność zmiany (adaptacja struktur społecznych, nowe idee, partie postępowe).
Po co nam w mózgu mapy topograficzne?
Mapy umożliwiają szczegółową analizę sygnałów zmysłowych.
Transformacje senso-motoryczne wymagają złożonych, nieliniowych przekształceń, mapy to umożliwiają.
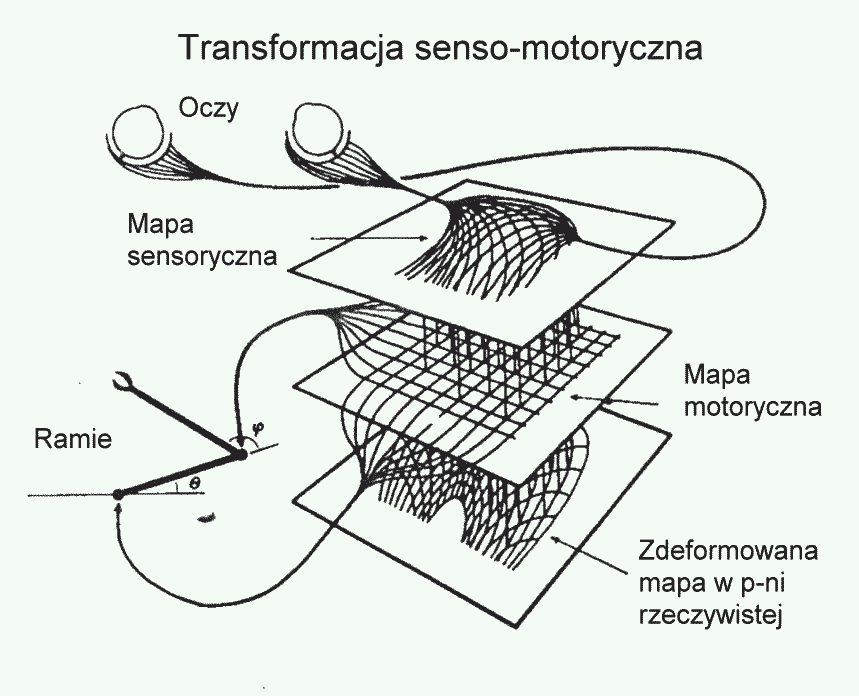
|
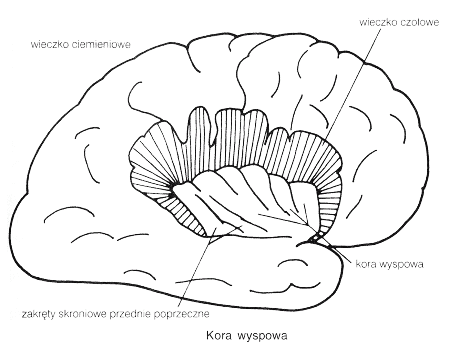
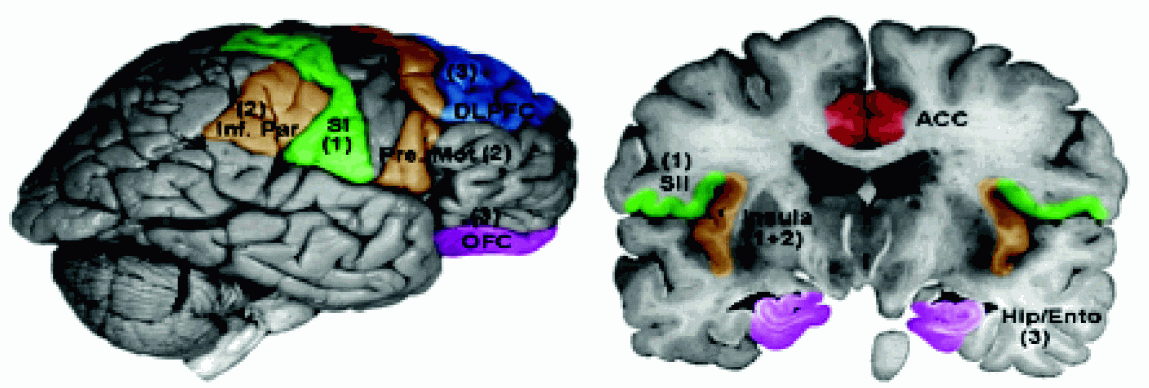
Wtórna kora czuciowa SII leży w głębi bruzdy środkowej i bruzdy bocznej (BA 43), w pobliżu kory wyspy, chociaż niektórzy zaliczają do SII również płacik ciemieniowy dolny (obszar BA5), nieco z tyłu w stosunku do SI.
Obszar ten wyróżniono tylko u ludzi i małp naczelnych, jest to obszar kojarzeniowy dla wrażeń dotyku i bólu. Komórki pobudzane są przez sygnały z większych obszarów ciała, z obu stron, bezpośrednio ze wzgórza, obszarów kory SI oraz spoidła wielkiego. Integracja informacji z obu stron ciała oraz hipokampu pozwala na uczenie się rozpoznawania przez dotyk i transfer tej umiejętności z jednej ręki do drugiej. | 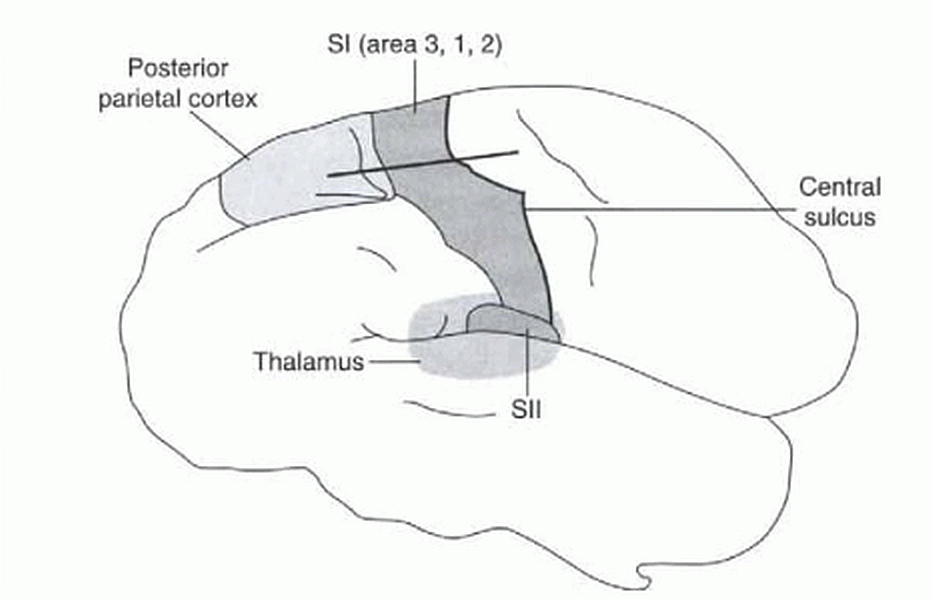 |
|
Uszkodzenia SII i pobliskiej kory wyspy powodują
asymbolię bólu,
brak cierpienia (reakcji psychicznych) na ból, lub wywołują
stały ból piekący (kauzalgia).
|  |
Damasio podaje opisy przypadków osób po operacji w okolicach kory zakrętu obręczy lub kory wyspy, po których pacjent nadal odczuwał ból, ale zniknęło cierpienie.
Środki przeciwbólowe obniżają aktywność kory przedniego zakrętu obręczy.
Ból wieńcowy spowodowany jest pobudzeniem tego samego obszaru.
Możliwa jest opóźniona reakcja na ból, reakcja następująca wiele minut po ukłuciu (tak jest w przypadku późnego stadium syfilisu).
Wtórna kora czuciowa SII zaangażowana jest również w interpretację emocjonalną sygnałów czuciowych.
Bodźce bólowe nie muszą przechodzić przez dokładniejszą interpretację na poziomie kory by wywołać mobilizację i reakcję organizmu.
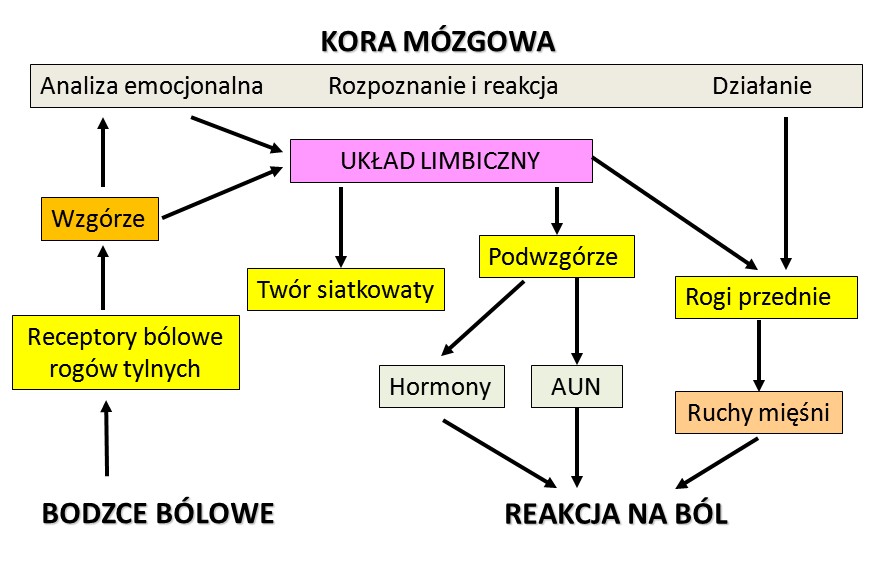
Uproszczony schemat dróg reakcji na ból.
SSA, dodatkowa kora czuciowa, znajduje się w płaciku ciemieniowym górnym (obszary 5 i 7 Brodmanna).
Obszary czuciowe w płacie ciemieniowym mają złożone funkcje:
- Stereognozja to identyfikacja dotykowa obiektów.
- Możliwe jest rozpoznawanie kształtów, rozmiarów i tekstur.
- Do utworzenia się zdolności sterognozji potrzebna jest nauka oparta na integracji wrażeń różnych modalności, szczególnie wzrokowo-czuciowych.
- Dermoleksja to zdolność do rozpoznawania kształtów (np. znaków) kreślonych na skórze np. palcem.
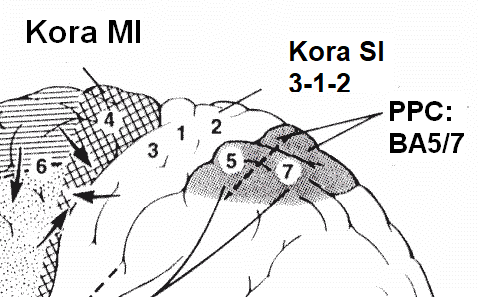
Kora czuciowa ma trochę neuronów kontrolujących ruch (tylko dwie synapsy dzielą ją od kontroli mięśni), a kora ruchowa neuronów czuciowych.
Czucie zewnętrzne to analiza sygnałów z narządów zmysłów.
Czucie głębokie to analiza sygnałów proprioceptywnych z mięśni, stawów, ścięgien, błędnika, informujące o położeniu i stanie ciała.
Czucie wewnętrzne, interoceptywne, analizuje informację z gruczołów i trzewi.
Agnozja (gr. a=negatywne, gnosis=wiedza) to niezdolność lub zaburzenie rozpoznania lub identyfikacji informacji zmysłowej.
Astereognozja to niezdolność do rozpoznania przedmiotów za pomocą dotyku.
Jest niezależna od zaburzeń czucia, upośledzenie rozpoznawania kształtu przy prawidłowym rozpoznawaniu wielkości, faktury.
Większość agnozji należy rozpatrywać na wyższym poziomie analizy powyżej kory analizującej dane zmysłowe.
Optyczna ataksja to niezdolność do chwytania przedmiotów pomimo prawidłowej analizy wzrokowej, np. nie można wlać wody do szklanki.
Uszkodzenia obszarów leżących w głębi przyśrodkowej kory ciemieniowej (LIP) wywołuje niezdolność do chwytania przedmiotów, np. lecącej piłeczki.
Obszar LIP zawiera topograficzną mapę obiektów istotnych z punktu widzenia bieżących możliwości działania, odwołania do specyficznych ruchów ciała i oczu, kategoryzację obiektów z punktu widzenia możliwych działań, spodziewane nagrody. LIP odpowiada więc za integrację informacji wzrokowo-przestrzennych, poznawczych i ruchowych, z mechanizmami uwagi.
Połączenie kory SI i SSA umożliwiają przepływ informacji pomiędzy lokalizacją wzrokową, a pobudzeniem czuciowym.
Przykłady:
Utrata propriocepcji w wyniku zapalenia nerwów prowadzi do ślepoty ciała!
Opis takiego przypadku jest w rozdziale "Bezcielesna kobieta" Olivera Sacksa, w książce
"Mężczyzna, który pomylił swoją żonę z kapeluszem".
Wystarczy informacja wzrokowa by kontrolować ciało, jest to możliwe dzięki połączeniu obszaru PPC 5 w korze ciemieniowej z korą czuciową SI.
Informacja wzrokowa może również spowodować, że nie zauważamy astereognozji.
Inny znany przypadek ślepoty ciała dotyczy Iana Watermana, pokazano go w filmie
"Człowiek, który stracił ciało" (krótka wersja
Ian Waterman's Missing Body. Długotrwały trening pozwolił mu na utworzenie wewnętrznej mapy ciała i na tyle precyzyjną wizualizację by częściowo przywrócić normalne funkcje kontroli ciała.
Na szczęście zaburzenia propriocepcji są rzadkie. Do 2020 roku badano zaledwie 5 przypadków osób z takimi zaburzeniami. Nie potrafią poruszać swoim ciałem nie widząc swoich członków. Z zamkniętymi oczami wykonują przypadkowe ruchy.
Jeszcze gorszym zaburzeniem jest ciągłe pobudzanie nerwów niosących informacje proprioceptywne. Może to powodować zaburzenia równowagi, wrażenie ciągłego spadania. Jedną z form terapii w tym przypadku jest substytucja zmysłów, dostarczanie informacji o położeniu ciała do mózgu np. przez nakładki na język. Pobudzanie nerwów, np. przez wibracje o określonych charakterystykach, wywołują wrażenie ruchu. Wibracje można tak dobrać by do mózgu dochodziły informacje podobne do tych, jakie są przesyłane w czasie wykonywania skomplikowanych sekwencji ruchów.
Kończyny fantomowe są wynikiem silnego odczucia obecności amputowanych kończyn, pojawiające się u 60-80% osób po amputacji.
Często jest to odczucie bolesne, czasami mogą się pojawić wszelkie inne wrażenia czuciowe: ciepło/zimno, swędzenie, ucisk, łaskotanie, wrażenie ruchu i próba chwytania nieistniejącą kończyną.
Odpowiedzialna jest za to reorganizacja kory czuciowej wynikająca z pobudzeń z okolic ciała, które mają zbliżone pola projekcyjne w obszarze SI, słabe połączenia normalnie maskowane przez odczucia płynące z brakującej części ciała.
Takie odczucia nie pojawiają się w przypadku porażenia kończyn dolnych (paraplegii).
Swędzenie i ból to całkiem odmienne odczucia, różne receptory i aksony neuronów (nerwy), różniące się szybkością przewodzenia impulsów.
Swędzenie można zredukować
pobudzając receptory bólowe przez tarcie, nacisk, ciepło czy drażnienie prądem. Drapanie blokuje przesyłanie sygnałów o swędzeniu przez rdzeń kręgowy. Drapanie może usunąć pasożyty lub czynniki chemiczne wywołujące swędzenie, a powstałe zadrapania
powodują stan zapalny zwalczający szkodliwe czynniki na skórze.
Bóle fantomowe pojawić się mogą u osób po amputacji (mają je prawie wszyscy krótko po imputacji, ale po dłuższym czasie zanikają, ma je tylko 2-4% osób), z uszkodzonym rdzeniem, z wrodzonym paraliżem.
Znany jest
przypadek osoby, która odczuwa fantom ruszający pięcioma palcami, chociaż urodziła się bez jednego palca, z dwoma innymi sparaliżowanymi, a rękę amputowano jej przed 35 laty.
Interpretacja tego przypadku nie jest całkiem pewna, ale nasuwa się sugestia, że obraz ciała jest w znacznej mierze genetycznie ustalony.
Znany jest też jedne przypadek osoby, twierdzącej, że widzi swoją kończynę fantomową.
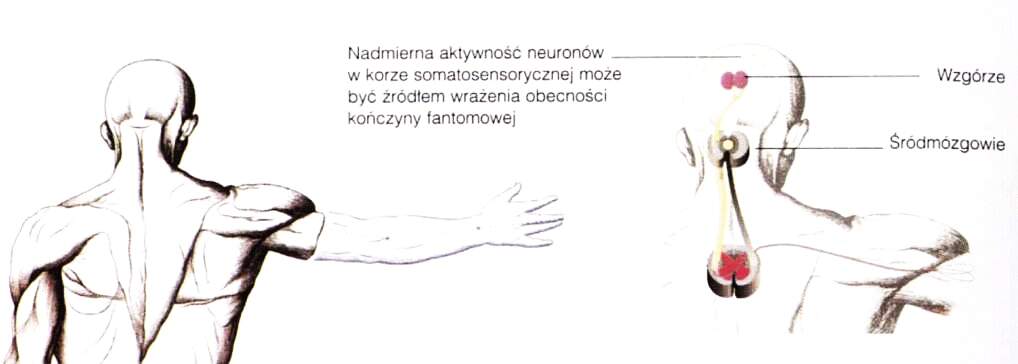
V.S. Ramachandran napisał książkę na temat chorych cierpiących z powodu kończyn fantomowych. Niektórzy cierpią na silne bóle, np. odczuwają wrastanie paznokci w zaciśniętą dłoń.
Ból wywołuje stymulacja obszarów, których reprezentacje przylegają do kory odpowiedzialnej za wrażenia z brakującej części ciała; prawdopodobnie wtórna kora SII dostaje mylne sygnały.
Możliwa jest też spontaniczna aktywność neuronów w wyniku uszkodzenia kory na skutek jakiejś lezji.
Terapia jest możliwa przez pokazywanie im ruchu własnych zdrowych kończyn,
odbitych w lustrze lub obserwowanie ruchu czyjejś ręki.
Informacja wzrokowa pobudza tu korę SI w obszarze odpowiadającym utraconej kończynie i wywołuje reorganizację tej kory, bez takiej informacji nie dochodzi do reorganizacji.
Obraz ciała w naszych mózgach jest w znacznej mierze wrodzony.
Ramachandran i Geoch (2007) opisali zjawisko fantomowych części ciała w przypadku narządów płciowych. Około 60% mężczyzn po amputacji penisa doświadcza fantomu. Być może podczas rozwoju embrionalnego mózgi osób transseksualnych są niewłaściwie "zakodowane", obraz ich ciała nie pasuje do płci biologicznej. W tej publikacji postawili hipotezy dotyczące doświadczania fantomowych narządów płciowych po amputacji u osób transpłciowych. To ciekawe hipotezy, ale osób transpłciowych po amputacji jest niewiele i trudno ją zweryfikować.
Eksperyment z butem i sztuczną ręką:
- ręka za zasłoną jest lekko uderzana jednocześnie z uderzeniami w but;
- korelacja powoduje poczucie, że but jest częścią ciała;
- silne uderzenie młotka w but wywołuje reakcję bólu.
Zrób sobie fantomową kończynę ...
Obejrzyj film z gumową ręką (Olaf Blanke, YouTube) i zajrzyj do laboratorium Ehrssona. Eksperyment z gumową ręką demonstruje też magik Justin Willman.
Symulacje posiadania ciała iluzorycznej osoby powstające w wyniku stymulacji elektrycznej obszaru TPJ (połączenia płatu skroniowego i ciemieniowego na końcu bruzdy Sylwiusza).
Dużo rzadszy przypadek to nadliczbowa kończyna fantomowa, której co prawda nie można użyć do manipulacji fizycznymi przedmiotami, ale można się nią podrapać! (Khateb i inn. 2009).
Eksperyment: Siedzimy na krześle z zawiązanymi oczami, a ktoś przed nami z przodu.
Wyciągamy dłoń i kładziemy ją na nosie osoby przed nami, a drugą dłoń kładziemy na swój nos i zaczynamy gładzić oba nosy. Do obu stron mózgu napływa więc informacja o gładzeniu nosa. Zawiązane oczy powodują, że nie wiemy czy to tylko nasz nos.
W ciągu niecałej minuty pojawia się odczucie posiadania całkiem sporego nosa. Czasami trzeba powtórzyć eksperyment kilka razy by pojawiły się takie odczucia.
Warianty: głaskanie nosa skrzyżowanymi palcami - czy mamy dwa nosy?
Głaskanie nosa manekina. Czy też będzie efekt?
Mamy obecnie dokładniejsze badania zjawisk dysocjacyjnych, analizujące powstawanie rytmów o niskiej częstotliwości 1-3 Hz w korze retrospenialnej (kora przylegająca do tylnej części płata spoidła wielkiego). Ta część kory jest istotna w postrzeganiu relacji przestrzennych, wyobraźni i pamięci przestrzennej. Badania robiono na pacjentach cierpiących na padaczkę z implementowanymi elektrodami, jak i na poziomie komórkowym i molekularnym na myszach.
Kończyny niechciane (dysmorfia ciała). Silny wstręt do własnego ciała, poczucie obcości, zdarza się dość często, takie odczucia ma 1:50 osób. Skrajne przypadki zaczynają się w dzieciństwie, "życie w ciele, które nie jest własne". Takie zaburzenie traktowane jest jako obsesja, niektórzy pacjenci walczą o prawo do amputacji kończyn (apotemnofilia). Kilku osobom w Anglii obcięto kończyny (BBC Horizon 'Complete Obsession').
Cenestezja to odczucia i wrażenia odbierane z wnętrza ciała.
Kończyny fantomowe są specyficznym rodzajem halucynacji związanych z pobudzeniem kory czuciowej, ale mogą się oczywiście zdarzyć różne halucynacje, np. cenestezyjne halucynacje oralne: wrażenia gazu wydobywającego się z ust, drutów albo ślimaka, który jest na dziąsłach.
Wrażenia, odczucia to interpretacja stanów różnych obszarów mózgu, przetwarzających dane zmysłowe.
Co może być przyczyną takich halucynacji czuciowych?
Sprzężenie obszarów interpretujących informację z kończyn w obszarze SII z ciałem migdałowatym może wywołać nieprzyjemne odczucia, możliwe są też inne odczucia, np. podniecenie seksualne.
Nie ma tu dobrych metod leczenia. Można spróbować pobudzania nerwów czuciowych w połączeniu z przyjemnymi bodźcami, co daje czasem dobre rezultaty.
Gdzie jestem? Skąd to wiem?
Nauczyłem się korelować wrażenia wzrokowe i czuciowe, wzrok dostarcza nam większość informacji do kory ciemieniowej, tworząc egocentryczną i allocentryczną mapę przestrzeni. Wyjaśnia to film Henrika Ehrssona z TEDx:
What if we could leave our body and have a new one?
Tworzenie obrazu ciała jest powolnym procesem. Dziecko nie trafia do swojej buzi nie tylko z powodu słabej kontroli ruchu kończyn, ale braku obrazu swojego ciała. Dopiero około 23 roku życia ten obraz się w pełni stabilizuje.
Inne obszary zorganizowane topograficznie, w których pojawia się informacja somatosensoryczna:
- móżdżek,
- jądro siatkowate wzgórza (NRT),
- wzgórki czworacze górne,
- inne jądra.
Modele przewidując ruch ciała na podstawie oceny parametrów wewnętrznych pokazują, że dobre rezultaty można osiągnąć korzystając z reprezentacji informacji o propriocepcji. Rozpoznawanie obiektów i afordancje, czyli możliwości oddziaływania na różne obiekty, związane są z propriocepcją i reprezentują różne typy obiektów fizycznych. Mózg musi przewidywać na podstawie takiej informacji, jest to więc istotna część reprezentacji pomagającej kontrolować ciało. Model obliczeniowy z pracy "Uczenie się świadomości działania" (Amoset i inn. 2018) pokazuje, że można przewidywać ruchy ciała tylko na podstawie sensorów zbierających informację o propriocepcji.
Nie zawsze uświadamiamy sobie sygnały zmysłowe. Słyszymy, ale nie reagujemy, bo skupiamy się nad czymś innym, obserwacją bądź myśleniem, więc to do nas nie dociera. Jest to związane z mechanizmami uwagi. Sygnały ze zmysłów wędrują
do kory przez wzgórze,
którego powierzchnia podzielona jest na wiele podobszarów, łączących się z różnymi nerwami.
Zmysły pobudzając różne części wzgórza: wzrok LGN, słuch MGB a czucie VB. Wokół wzgórza rozciąga się
jądro siatkowate wzgórze (jądro RTN, nie należy je mylić z tworem siatkowatym w pniu mózgu), które łączy różne obszary i dzięki pętlom wzgórzowo-korowym może zmniejszając aktywność niektórych z nich, osłabiając wpływ wybranych dróg zmysłowych na działanie kory. Jeśli kora - sieć określająca istotność bodźców - uzna w danej chwili pewne sygnały za mniej istotne może wpłynąć na jądro RTN i wyhamować dopływ informacji z nich płynący.
ASMR (Autonomous Sensory Meridian Response) to przyjemne mrowienie zaczynające się w okolicach głowy i zstępujące po szyi na ramiona i inne obszarach ciała, wywołane przez różne bodźce zewnętrzne, takie jak szepty, szelesty, wrażenia dotykowe.
Wygląda to na przykład
parastezji, mrowienia i drętwienia, które jest zwykle nieprzyjemne, ale ASMR wywołuje stan przyjemny, używany do relaksacji.
Nie każdy ma takie wrażenia, ale
wykonano eksperymenty
potwierdzające u osób odczuwających ASMR realny wpływ na stany emocjonalne, puls i GSR. Jest wiele filmów o ASMR, które warto obejrzeć by zbadać swoje własne reakcje.
Gęsia skórka, dreszcze emocjonalne odczuwane pod wpływem muzyki, zwane fachowo
frisson, to odmienne zjawisko. Gęsia skórka to pozostałość po wczesnym etapie ewolucji, stających włosach czy stroszeniu piór gdy robi się zimno lub zwierzę chciało wyglądać na większe, bardziej groźne.
Nie zawsze towarzyszy to uniesieniom emocjonalnym pod wpływem muzyki, ale muzyka pobudza bardzo różne procesy w mózgu, które odczuwamy jako emocje.
A14.4 Kodowanie populacyjne i kontrola ruchów
|
Oprócz analizy danych zmysłowych najważniejszym zadaniem mózgu jest kontrola działania, a więc ruchu ciała.
Mamy 640 mięśni szkieletowych; nawet najprostsze ruchy wymagają bardzo złożonej kontroli, której nie jesteśmy w pełni świadomi.
Zrobienie jednego kroku wymaga koordynacji większości mięśni.
Kwestie planowania działań należą do wyższych czynności poznawczych i będą omawiane później. Szlaki ruchowe biegną przez śródmózgowie (droga korowo-rdzeniowa) lub przez móżdżek do pnia mózgu:
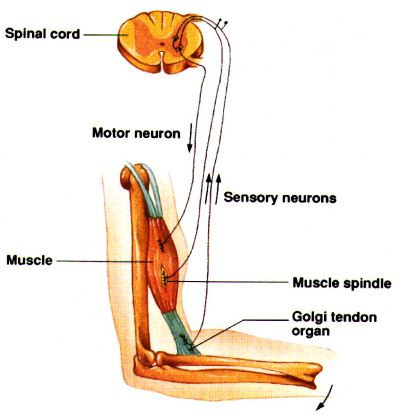
Lokalne odruchy rdzeniowe biegną od receptorów przez rdzeń do mięśni, tworząc pętle sprzężeń zwrotnych. Większość ruchów intencjonalnych wymaga sprzężenia postrzegania rezultatów działania i dostosowania sygnałów kory ruchowej by je skorygować. Dlatego mówimy o działaniach senso-motorycznych, sygnałach zwrotnych z propriocepcji, wzroku i zmysłu równowagi koniecznych do działania. Często powtarzane działania stają się automatyczne, zakodowane w pamięci mięśniowej. |
|
|
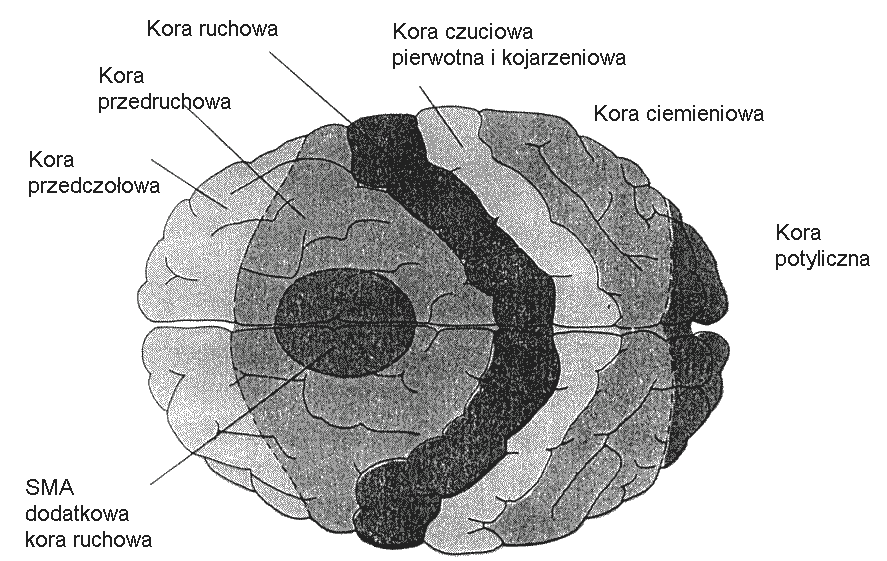
Mamy trzy główne obszary kory, kontrolujące ruch:
Dodatkowo czołowe pole okoruchowe (z przodu SMA) kontroluje wolicjonalne ruchy oczu. |
|
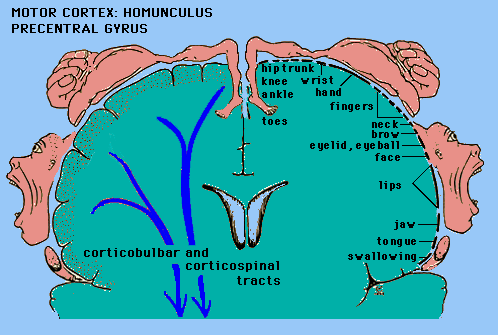
Pierwotna kora ruchowa (MI) ma organizację topograficzną (somatotopową), tworząc "homunculus motoryczny".
Pobudzenia tej kory wywołują precyzyjne ruchy, pojedyncze mięśnie/ścięgna (latencja 60-80 ms), ale bez poczucia chęci ruchu.
Zrób sobie mapę kory ruchowej (tylko lokalnie).
Mięśnie twarzy i głowy pobudzane są obustronnie, pozostałe przeciwlegle.
Pobudzenia dochodzą do MI z :
- kory czuciowej (bezpośrednia reakcje na bodźce);
- brzusznobocznego jądra wzgórza (połączonego silnie z móżdżkiem);
- kory przedruchowej MII (ruchy planowane).
Dodatkowa kora ruchowa (SMA) ma również częściowo organizację topograficzną.
Stymulacja wywołuje złożone ruchy, przygotowanie postawy ciała.
Jej rolą jest organizacja sekwencji ruchów, np. obu rąk, oraz działania ruchowe wymagające pamięci.
Kora przedruchowa MII jest 6 razy większa od kory MI, wykonuje bardziej złożone zadania.
Boczna kora przedruchowa odpowiedzialna jest za wolicjonalne działania senso-motoryczne, na podstawie danych zmysłowych.
Obrazowanie mózgu pokazuje aktywność MII podczas zadań wymagających koordynacji wzrokowo-ruchowych lub słuchowo-ruchowych.
Stymulacje wywołują stereotypowe ruchy złożone: kiwanie, obrót głowy lub tułowia.
Brzuszno-boczna kora przedruchowa aktywizuje się przy celowych aktach ruchowych, np. chwytaniu niezależnie od sposobu uchwytu, lewej lub prawej kończyny lub warg.
Lezje MII prowadzą do apraksji, niezdolności do wykonywania złożonych ruchów, np. chodzenia.
Jak to działa? Kodowanie populacyjne
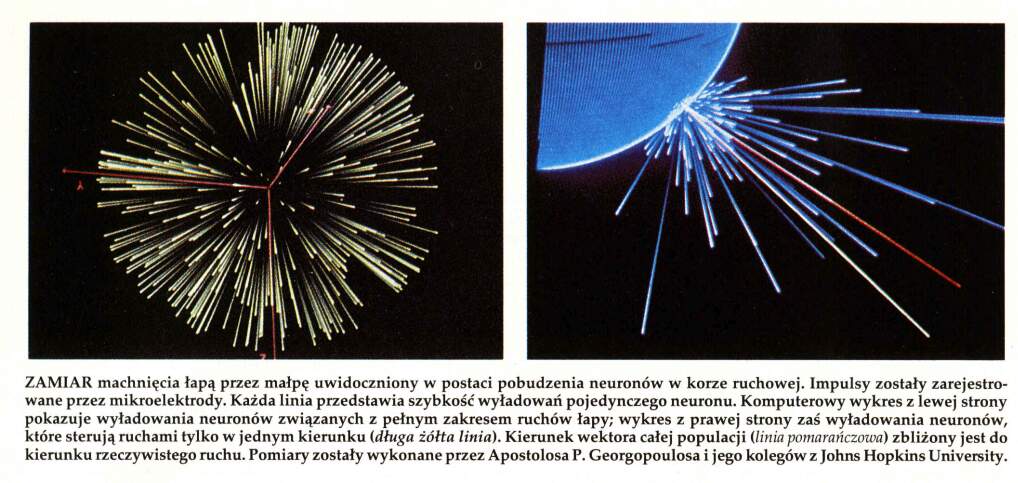
Rysunek pokazujący kierunki ruchu, przy których dana komórka najsilniej reaguje.
Dany neuron pobudza się najsilniej przy ruchu w określonym kierunku i słabiej przy ruchu w podobnych kierunkach, a prawie wcale przy innych ruchach.
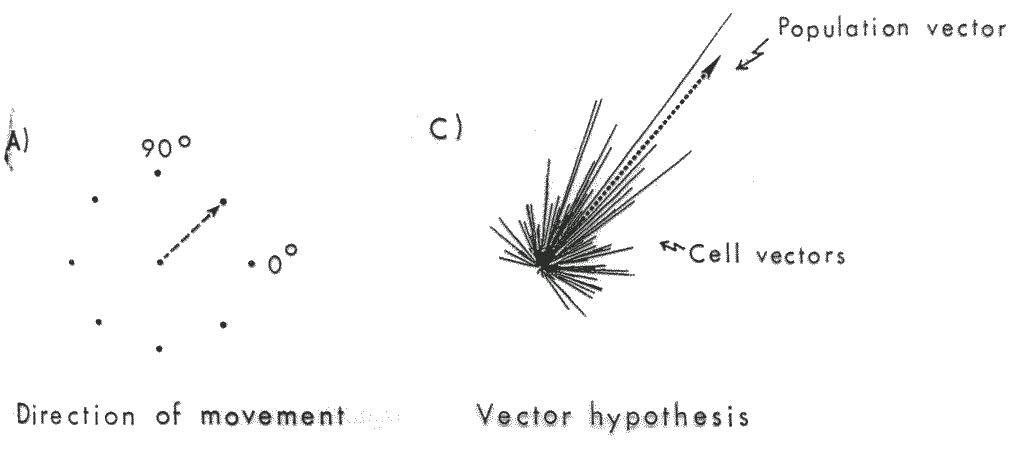
Długość strzałki zależy od częstości impulsacji neuronu w danej populacji, wzbudzającego się przy ruchu w danym kierunku.
Już dla kilkuset aktywnych komórek sumowanie daje najsilniejsze średnie pobudzenie zgodne z prawdziwym kierunkiem ruchu.
Najsilniej reagujące komórki są w MI, kolumna o średnicy ok. 1 mm.

W korze ruchowej decyzje podejmowane są przez uśrednioną aktywację całej populacji, nie przez pojedyncze neurony.
Kodowanie populacyjne: w MI, MII, SMA, różnych obszarach płatów kory i móżdżku.
Dekompozycja złożonych pobudzeń na pobudzenia elementarne: lokalne funkcje bazowe.
Można zaobserwować przejście od ruchu palcami równolegle do siebie, do ruchu nożycowego, zależnie od częstotliwości metronomu: miliony neuronów muszą działać synchronicznie i robią to tylko na dwa sposoby.
Do pewnego stopnia – chociaż znacznie mniej dramatyczne – pokazują to proste eksperymenty. Np. siedząc podnosimy prawą nogę i kreślimy stopą koła zgodnie ze wskazówkami zegara, a po zrobieniu 10 ruchów próbujemy napisać prawą ręką cyfrę 6 - co się dzieje z nogą?
Terapie ruchowe: są dane świadczące o wzmożonej neurogenezie na skutek ruchu; mówi się o "somatycznej edukacji".
Do terapii ruchowych należy
metoda Feldenkreisa, zakładającej, że świadoma percepcja szczegółów ruchu (metoda Awareness Through Movement) pomaga w jego doskonaleniu i rehabilitacji.
Nawet zwykłe
szybkie chodzenie
pół godzinny dziennie pozytywnie oddziałuje na poprawę nastroju, zmniejsza ryzyko depresji, zwiększa przepływ krwi do mózgu poprawiając pamięć, a nawet kreatywność i różne funkcje poznawcze. Ostatnie badania pokazują też, że składnia języka mówionego wykorzystuje jądra podstawy mózgu, podobne struktury neuronowe jakie służą kontroli złożonych ruchów.
Mowa gestów: u ludzi czarnoskórych dłonie są jaśniejsze, co prawdopodobnie pozwala na lepszy kontrast wzrokowy w czasie gestykulacji.
Plemiona afrykańskie i inne społeczności zbieracko-myśliwskie w czasie opowiadań używają gestów podobnie jak w pantomimie, naśladując zwierzęta i ludzi.
Szympansy na wolności rozpoznają przynajmniej 66 różnych gestów, służących im do komunikacji (Hobaiter, Byrne , 2011).
Osoby głuche używają
języka migowego,
korzystają przy tym z tych samych obszarów co w przypadku rozumienia i produkcji mowy.
Jest kilka języków migowych, dość znacznie się od siebie różniących, w tym
Polski Język Migowy oraz odrębny
(i często z nim mylony)
System Językowo-Migowy, służący do wspomagania wypowiedzi dźwiękowych znakami migowymi.
Poczucie rytmu i zdolność do poruszania się w rytm słyszanych dźwięków jest głównie cechą człowieka, nie potrafią tego zwierzęta.
Chociaż sygnały kontrolujące ruch członków po jednej stronie ciała pochodzą od kory ruchowej o przeciwległej stronie okazuje się, że intencje ruchu można dostrzec w korze ruchowej po tej samej stronie. W przypadku udarów mózgu udało się odzyskać cześć funkcji motorycznych dzięki nieinwazyjnej stymulacji mózgu za pomocą interfejsu mózg-komputer.
Ciekawostka: dlaczego kura bez głowy biega?
Zwykle bez głowy można biegać tylko krótko, ale kogut Mike przeżył 18 miesięcy! Miał nieuszkodzony pień mózgu, właściciel odżywiał go kroplówkami i pokazywał za pieniądze.
Graficzne podsumowanie (text2mindmap).
{kind=link}
W następnej części omówimy dokładniej percpecję wizualną i jej relacje z mechanizmami kontroli ruchu.
Pytania, na które powinniście znać odpowiedzi po przeczytaniu notatek do tego wykładu:
- Ogólne informacje: wielkość kory mózgu, typy kory.
- Budowa laminarna i kolumnowa kory - jakie kolumny, jak duże, ile ich jest, jaka jest ich rola?
- Czy kora jest niezbędna do inteligencji?
- Co to jest kranioskopia, frenologia, fizjognomika, psychognomika?
- Czym są pęczki, obręcze, spoidła, projekcje aferentne/eferentne?
- Jakie są możliwości lokalizacji funkcji psychicznych?
- Wymienić 3 najważniejsze funkcje kory mózgu.
- Jakie mamy pierwotne obszary zmysłowe? Jakie wtórne? Jaka jest ich rola?
- Dlaczego najszybsze czasy reakcji mamy około 40 roku życia?
- Co to jest hydranencefalia?
- Jakie ograniczenia ma neuroplastyczność i czym to jest uwarunkowane ?
- Czy nie mając kory mózgowej można normalnie funkcjonować?
- Czym różni się neurologia, psychiatria i psychologia kliniczna?
- Dlaczego badania konektomu są takie ważne?
- Gdzie znajduje się pierwotna kora czuciowa i jakie informacje otrzymuje?
- Jaka jest rola somatosensorycznych map topograficznych?
- Gdzie jest wtórna kora czuciowa i jaką pełni role?
- Gdzie leży dodatkowa kora czuciowa i jaka jest jej rola?
- Co to jest sterognozja? Dermoleksja? Ślepota ciała?
- Skąd się biorą bóle fantomowe?
- Dlaczego mózg Einsteina nic nam nie mówi na temat inteligencji?
- Jakie obszary mózgu zaangażowane są w kontrolę ruchu?
- Jaki procent wszystkich neuronów mamy w móżdżku? Dlaczego jego uszkodzenia nie powodują upośledzenia funkcji poznawczych?
- Co to są mapy ruchowe i w jakiej części mózgu je znajdziemy?
- Na czym polega i do czego służy kodowanie populacyjne.
- Wymienić główne funkcje kory przedruchowej.
- Co to jest stereognozja i jaka część mózgu jest za nią odpowiedzialna?